
What Is Blue Light?
Human eyes have evolved to function in the environmental conditions of Earth: a continuous cycle of sunlight and darkness. However, the development of artificial light has drastically changed our environment and created a number of challenges for our visual system: not only has it affected the patterns of light and dark that govern our circadian rhythms, it has also changed the nature of light to which our eyes are exposed.
The widespread use of light-emitting diode (LED) lighting as a screen lighting component in computers, smartphones, tablets, and TVs has been the latest hurdle for our visual system. The problem with LEDs used in screens is that they emit disproportionate amounts of blue light that’s stressful to the retina. And with the amount of time people of all ages are spending in front of screens, blue light exposure is becoming a very significant cause of stress to the eyes for much of the population. Heavy use of digital devices has been linked to a number of symptoms of eye strain and visual fatigue. Some of these symptoms are partly the result of prolonged exposure to blue light.
Let’s explore how blue light exposure may affect our eyes and how the eyes protect themselves from damage.
How Can Visible Light Be Stressful To The Eyes?
Visible light is what we actually see—the light that is emitted or reflected by the things that surround us and is captured by our eyes. [If you wish to learn about light, eye structure, and how vision works, check out our overview of the visual system.]
Only visible light is able to reach the retina (the sensory tissue of the eyes) and be captured by photoreceptors (the sensory neurons of the eyes) to generate visual stimuli. There is some infrared (IR) radiation that reaches the retina, but it is not visible. Most electromagnetic radiation outside of the visible spectrum is filtered as it passes through the cornea, iris, lens, and vitreous [1]. This ability of eye tissues to filter light is important because it keeps radiation with too much light energy, particularly ultraviolet (UV) radiation, away from the highly sensitive retina (as long as you don’t look directly at the sun). However, the visible light that reaches the retina does contain high-energy radiation that is not totally harmless.

Figure 1 - How eye tissues filter electromagnetic radiation. Source (adapted): Iliya V. Ivanov et al. 2018. Licence CC BY 4.0
The eye has a number of molecules with the ability to absorb light. They do so because they contain in their structure a chemical moiety called chromophore—from the Greek words for “color” and “carrier” because this is the part of molecules responsible for giving color—that is able to capture the energy of light particles (i.e., photons). Pigments, like those found in the iris that give the eye its color, or those in the macula area of the retina that give that part of the eye a yellow appearance, are examples of molecules that absorb light. Photoreceptors in the retina have pigments that absorb specific wavelengths of light as part of the visual process, allowing us to see colors and in dim light, and to signal circadian rhythms. But, while absorbing light energy provides lots of benefits, it also comes with a cost; the eye needs to protect itself against the stress caused from absorbing light [2,3].
The eyes evolved to capture light energy… and to protect themselves from it. Three macular pigments—lutein, zeaxanthin, and meso-zeaxanthin—absorb blue light. They are a part of the protection mechanisms.
The absorption of light energy by some pigments may result in the generation of heat—radiant energy from light is transferred into the pigments absorbing it in a way that is not too dissimilar to a sidewalk or car seat warming up from sunlight. That’s the case with melanin in the retinal pigment epithelium and vascular layer, for example (melanin is also the pigment that gives eyes their color and is involved in tanning from sunshine). When exposed to intense light, these pigments may generate heat at a rate that overpowers the eye’s ability to dissipate it via blood flow (blood flow is acting like a cooling system), causing a rise in the temperature of tissues in the eye. This is known as photothermal stress. It is a type of light stress that occurs because radiant energy from light is being absorbed at a faster rate than the retina of the eye can cool itself off.
Increased temperature above a critical amount is stressful because it can cause the denaturation of proteins (i.e. the loss of their 3D structure) and, consequently, inactivate them, leading to tissue dysfunction. But photothermal stress doesn't happen easily: it would require a temperature rise of around 16°F/10°C, which is way above the average rise of 3.6°F/2°C of a human retina exposed to direct sunlight. However, this is the case mostly because the pupil constricts fully to protect the eye; with a dilated pupil, not only would photothermal injury to the retina be possible, it would develop in only a few seconds [2,3].
[Note: The ‘sunburn’ of the eyes caused by looking directly at the sun is not actually caused by heat, it’s caused by a metabolic overload of photoreceptors and by the action of potentially harmful metabolic byproducts that result from the intense stimulation of photoreceptors by light. (More detail below.)]
Protecting themselves from photothermal stress is not the only type of heat issue eyes have to worry about. If a lot of light energy is introduced in a very brief period of time, not only can it increase temperature by more than 16°F/10°C, the heat can also produce a thermoelastic pressure wave (think of this as being akin to a shock wave rippling through the retina) because tissue expansion is occurring faster than mechanical relaxation can diffuse it. The result is mechanical stress due to compressive or tensile forces, which can lead to substantial tissue injury. This is called photomechanical stress and it requires very high local temperatures. It’s the type of light toxicity that can be caused by lasers, for example.
There’s another type of light toxicity that occurs without changes in tissue temperature. It results from the generation of potentially harmful chemical compounds in cells that are produced as a side-effect of light absorption. This is called photochemical stress. It develops more slowly and requires longer exposures to the light source (with the duration of exposure needed to induce stress being dependent on the intensity of the light source) [2,3].
[Note: The type of retinal injury you get if you look directly at the sun is a result of photochemical stress. Even with fully constricted pupils, the activation of photoreceptors by light is so intense that there is a peak in the production of potentially harmful chemical compounds as byproducts of the metabolic overload photoreceptors are being subjected to. You’re unable to see for a while because photoreceptors are saturated and visual pigments have all been quickly used up; as they are recycled, you regain vision. The phototoxic effects of staring at the sun then develop gradually over the course of a few hours.]
And whereas photothermal light toxicity can be caused by all wavelengths of light within the visual spectrum, i.e., there is no color that is noticeably more harmful, the same isn’t true for photochemical stress: blue light is the biggest culprit [4].
When we spend hours looking at screens, we are exposing our eyes to ‘photochemical’ stress, a type of light stress that occurs because of the chemical reactions and oxidative stress from the retina absorbing blue light for prolonged periods of time.
Blue light corresponds to the shorter wavelength end of the visible light spectrum, ranging from around 380 to 500 nm. Wavelength is inversely correlated with the energy of the light rays, which means that the blue end of the spectrum has higher energy than light towards the other end of the spectrum, i.e. red light. That’s why eyes, in general, and the retina and its macula, in particular, have evolved mechanisms to protect themselves from blue light.
[Note: This wavelength interval is called blue light but it also includes violet light (think of the colors of the rainbow). It is sometimes split into two categories: blue-violet light (around 380-440 nm) and blue-turquoise light (around 440-500 nm).]
The type of photochemical stress to the retina caused by blue light is known as blue light hazard or retinal phototoxicity [5]. The harmful effects of blue light (including LED light) to retinal cells have been studied for decades and demonstrated in many studies [5–9].
The degree of phototoxicity blue light can cause is dependent on a number of factors: the intensity of blue light to which the eye is exposed, the distance to the source of light, the direction of the line of sight, and the spectrum of the light source, for example. It’s important to clarify that the main source of blue light in our environment is, without doubt, the sun. But although the amount of blue light emitted by a screen is low compared to sunlight, the fact is that the type of exposure is very different. Anyone who spends long periods of time looking at screens, especially in close proximity, is being continually exposed to a significant amount of blue light that’s different in important ways compared to looking at a blue sky or ocean.
Mechanisms of Blue Light-Induced Retinal Stress
The agents of photochemical light stress are reactive oxygen species (ROS), a type of molecule that contains oxygen and that, due to its chemical instability, reacts easily with other molecules in a cell to oxidize them. ROS are naturally produced in cells, where they have important functions as signaling molecules. Because the main job of the eyes is to capture light, and ROS are a metabolic byproduct of this work, cells in the eye have antioxidant defenses to neutralize ROS. But when ROS production is unbalanced, exceeding the ability to neutralize them, cells fall into a state known as oxidative stress that can be harmful to cell function [10].
ROS can be produced by photosensitizing compounds in the retina. These are pigmented molecules in photoreceptors that absorb light energy (from photons, light particles) and then transfer that energy to neighboring molecules (as electrons, negatively charged particles), generating damaging versions of those molecules (free radicals, highly reactive molecules). When photosensitizers transfer energy to oxygen (O2), ROS are formed.
This means that, when activated by light, photosensitizers generate ROS. Because the retina has many potential photosensitizer molecules and because it is so exposed to light and oxygen, the potential for generation of ROS by photosensitizers in the retina is particularly high [3]. This is why antioxidant defenses are such a critical part of retinal health.
Among the main blue light-absorbing photosensitizers in the retina are molecules of the visual pigments of photoreceptors (retinal and opsins) and molecules of the mitochondrial electron transport chain (flavin and porphyrin-containing enzymes). The latter are particularly relevant for blue light phototoxicity.
One of the main reasons that blue light is stressful to the eyes is that the work of capturing and using light produces oxidative stress. This is why antioxidants are so important for eye health, and even more important when there’s lots of blue light exposure.
The Role of Mitochondria in Blue Light Stress
Mitochondria are the cellular organelles responsible for the production of cellular energy in the form of ATP. The mitochondrial electron transport chain (ETC) is a chain of enzyme complexes that use the energy of electron transfers to power the production of ATP in a process known as oxidative phosphorylation. The ETC is also a major site of ROS production as a byproduct of energy metabolism, due to the leakage of electrons from the ETC [11,12].
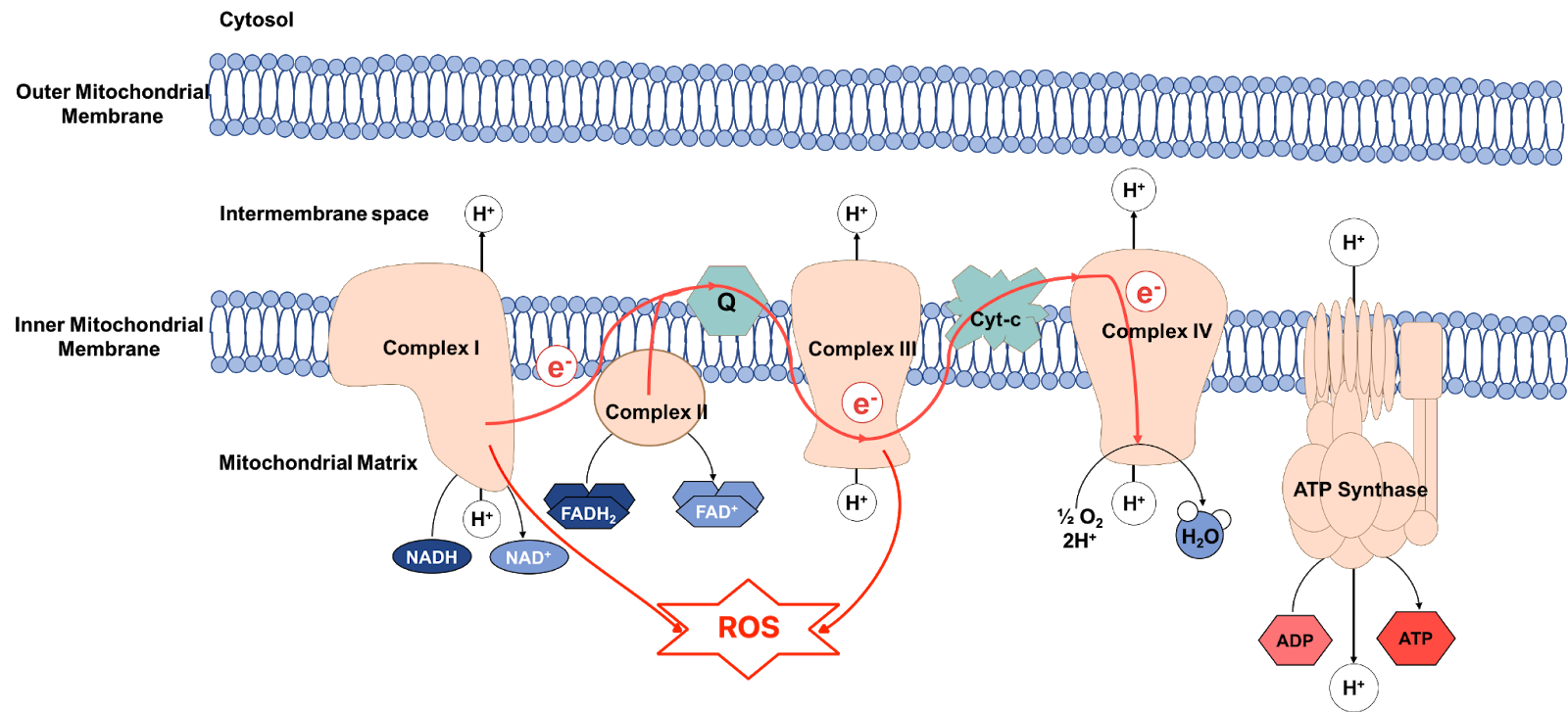
Figure 2 - Mitochondrial electron transport chain. Source (adapted): Dw001 Wikimedia Commons. Licence: CC BY-SA 4.0
Studies have shown that photosensitizers in the retina have a negative impact on the structure and function of mitochondria and that blue light can interact with neuronal mitochondria to cause a reduction in energy levels and an increase in the generation of ROS [12].
Blue-light induced ROS production can lead to a state of photooxidative stress that results in the oxidative damage of cellular membranes. This has a significant negative impact on mitochondrial function and ETC efficiency because the ETC is located at the inner mitochondrial membrane. And loss of ETC efficiency increases the generation of ROS. So, we’re looking at a vicious circle in which ROS stress mitochondria and stressed mitochondria produce more ROS [11,12].
Blue light-induced ROS production can also stress DNA in retinal neurons and epithelial cells, both nuclear (nDNA) and mitochondrial DNA (mtDNA). But because mtDNA is less protected and has a less efficient repair system than that of nDNA, mtDNA is more susceptible to oxidative stress. Damaged mtDNA further disrupts the integrity of the mitochondrial ETC, contributing to the vicious circle of ROS-mediated mitochondrial dysfunction [5,13–15].
As a result, mitochondrial activity further deteriorates leading to decreased ATP production, the activation of damaging signaling pathways, and the disruption of mitochondrial structure, eventually leading to irreversible loss of mitochondrial function. This triggers a number of unhealthy processes within cells of the retina: cytokine signaling cascades that lead to loss of blood-retinal barrier integrity, blood vessel damage, microglia activation, immune cell (macrophage) recruitment; ultimately, it may lead to death of retinal cells and tissue dysfunction [5,11,16–18].
Retinal mitochondria are both a major source and a major target of blue light-induced ROS. Because of their roles as mitochondrial antioxidants, ingredients like astaxanthin and bilberry play important roles in supporting eye health.
Retinal neurons, particularly photoreceptors, are particularly susceptible to blue light-induced mitochondrial dysfunction because they have large numbers of mitochondria to provide for their high energetic requirements. Similar to the brain, which consumes approximately 20% of the total oxygen used in the human body, the retina has massive oxygen demands for phototransduction and neurotransmission and its rate of oxidative metabolism is among the highest in the human body [12,19,20].

Figure 3 - Distribution of mitochondria (yellow dots) in the retina. Source (adapted): Jin-Xin Tao et al. 2019. Licence: CC BY 4.0
But blue light stress is not restricted to the retina; it also affects the cornea, the first barrier of the eye against light [21,22], for example. Blue light hazard on the ocular surface (the cornea and the conjunctiva) is also driven by excessive ROS production, mitochondrial dysfunction, damaging cytokine signaling, and macrophage recruitment [5,23]
How Aging Influences Susceptibility to Blue Light
The lens absorbs UV radiation (longer UVB and the full range of UVA, i.e., between 295 and 380 nm) and part of the near IR radiation. It uses protective yellow pigments that absorb and dissipate the energy from UV light. By preventing UV waves from penetrating deeper in the eye, this protects the more sensitive retina from the high energy carried in UV wavelengths. The lens is clear at birth and gradually develops the protective yellow pigments during the first few months of life. But the absorption spectrum of the lens changes with age for more than UV—the absorption of blue light gradually increases as we get older and the result is that less blue light reaches the retina. This means that young children’s eyes absorb more blue light than adult’s eyes and that the absorption of blue light keeps dropping with age [24–26].
This would seem to indicate that the risk of blue light damage decreases as we grow older, but it’s not that simple. As we age, there is also a gradual accumulation of lipofuscin in many biological tissues. Lipofuscin is a pigment composed of residues of cellular digestion that accumulates within lysosomes. Lysosomes are cellular organelles that (among other functions) participate in the disposal and recycling of cellular waste by digesting large molecules, old cell components, and extracellular structures . Lipofuscin accumulation is relevant because lipofuscin absorbs visible light and can act as a photosensitizer.
In the eye, lipofuscin accumulates most extensively in the retinal pigment epithelium (RPE) [27]. The RPE plays important parts in eye health by supporting photoreceptor function, providing metabolic support, and maintaining the structural integrity of the retina, for example. One of the functions of the RPE is to clean up parts of photoreceptor cells which undergo constant destruction due to photooxidative damage that’s a normal byproduct from capturing light energy [28–30].
Lipofuscin, through its component A2E, sensitizes the RPE to blue light photodamage. Under blue light stimulation, A2E stimulates ROS production, leading to mitochondrial dysfunction, inhibition of lysosomal autophagy by oxidative stress damage, and RPE cell death by apoptosis [31–34]. Given the important functions of the RPE, these changes can result in the gradual loss of retinal health.
So, overall, although there’s less blue light reaching the retina as we age, the light that does reach it is potentially more harmful due to the accumulation of lipofuscin.
Support against blue light is important for all ages: children because the lens filters out less blue light, so more of it can get to the retina; and adults because, while more is filtered by the lens, what does get to the retina can be more stressful.
How Does The Eye Protect Itself From Blue Light?
The eye is great at protecting itself from light energy. The fact that it filters out most wavelengths of electromagnetic (EM) radiation—visible light is a very narrow band of the overall EM spectrum—before they can penetrate deep into the eye, such as UVR, is a great testament to that capacity. But because blue light is part of visible light, the eye does not block it all out. Instead, the eye has a number of defenses that help mitigate the effects of blue light, most notably, macular pigments.
The macula is the region of the retina that surrounds the fovea, the central region of sharpest vision. It is a pigmented area because it accumulates high levels of carotenoid pigments, known as macular pigments. In fact, because these carotenoids give this region the appearance of a yellow spot, the macula is also known as macula lutea, which means yellow spot in Latin [35].
Macular pigments are composed mainly of three isomeric xanthophyll carotenoids: lutein, zeaxanthin, and meso-zeaxanthin. Lutein and zeaxanthin carotenoids must be obtained from the diet (they are generally found together in many plant sources) and accumulate in the macula; meso-zeaxanthin can be produced in the retina from lutein [36].
Macular pigments absorb light with peak absorption at 460 nm, corresponding to blue light [37]. One of the physiological functions of blue-light absorption by macular pigments is supporting visual performance by attenuating chromatic aberration, light scatter, veiling luminance, and blue haze induced by blue light [38–40]. But absorbing blue light has the added benefit of protecting against the stress caused by this type of light.
Macular carotenoids are also strong antioxidants, capable of scavenging ROS, reducing oxidative damage, and reducing lipofuscin formation [36,41–43]. Because of these properties, macular pigments act as optical filters for blue light and provide antioxidant protection to the retina, attenuate oxidative injury at the macula, and help to maintain the structural and functional integrity of biological membranes.
Macular pigments thus have a key role in supporting eye health [36,43–46]. Higher levels of macular pigments are linked to improved macular function, reduced discomfort associated with glare and improved photostress recovery time (the time necessary to recover vision following exposure to a bright light) [38,39].
Qualia Vision includes Lutemax® 2020 because it contains all three macular pigments—lutein, zeaxanthin, and meso-zeaxanthin. These pigments are what the macula relies on as its blue-light blockers.
How to protect eyes from blue light nutritionally?
Lutein and zeaxanthin must be obtained from the diet. It’s important to have adequate levels of macular pigments in the diet to ensure adequate biological blue light protection for our eyes.
However, it has been determined that the vast majority of Americans do not consume lutein and zeaxanthin in amounts sufficient to produce increases in macular pigment levels [47], which may increase their susceptibility to poor visual health. Several studies carried out throughout the world (including many in the U.S.) have shown that average MPOD in the population is, more often than not, at levels that provide inadequate photoreceptor protection [48]. But changes in dietary patterns [49] and/or supplementation with lutein, zeaxanthin, and meso-zeaxanthin [50] may lead to meaningful changes in macular pigment levels.
This means that supplementing macular carotenoids can be a good way of supporting visual health and ensuring that our eyes’ ability to protect themselves from blue light is at its best.
That’s why we have included macular carotenoids in Qualia Vision, to be released this June. Macular carotenoid supplementation, by supporting the levels of macular pigments in the eyes, has been shown to support retinal and visual function and to support resistance to photostress, eye strain, and visual fatigue [51–66].
But it's not just macular pigments that can support our eyes’ defenses against blue light. Qualia Vision includes other ingredients that provide nutritional support for blue light protection.
Astaxanthin, another carotenoid with potent antioxidant effects that can cross the blood-retinal barrier, supports the retina from light stress [67].
Bilberry extract, which is rich in antioxidant polyphenols, provides protective support against blue LED light-induced retinal photoreceptor cell stress through antioxidant defense pathways [68,69].
Goji fruit extract is rich in bioactive antioxidant compounds, including the blue light filtering zeaxanthin (it’s considered one of the richest food sources of this macular pigment), but also polysaccharides, believed to be responsible for many of Goji’s health benefits. Goji offers protective support from intense bright light for photoreceptor cells in the retina through the upregulation of antioxidant defenses [70,71].
Taurine, an amino acid found in all ocular tissues, supports retinal and photoreceptor cell function and supports their protection against blue light-induced photooxidative stress [72,73].
The spice saffron is rich in bioactive compounds among which are zeaxanthin and other carotenoids, such as crocin, responsible for saffron's color. Saffron extract protects retinal cells against light-induced stress by supporting antioxidant protection and by helping to maintain their morphology and function [74–77].
Because of the high amounts of blue-light stress produced by devices with screens, Qualia Vision was formulated to offer layers of blue light protection by combining Lutemax® 2020, MirtoSelect® Bilberry Fruit Extract, AstaPure® Extract as a source of Astaxanthin, Saffron Stigma Extract, Goji Fruit Juice Extract, and Taurine.
References
[1]D.H. Sliney, Int. J. Toxicol. 21 (2002) 501–509.
[2]D. van Norren, J.J. Vos, Eye 30 (2016) 169–172.
[3]R.D. Glickman, Int. J. Toxicol. 21 (2002) 473–490.
[4]M. Marie, K. Bigot, C. Angebault, C. Barrau, P. Gondouin, D. Pagan, S. Fouquet, T. Villette, J.-A. Sahel, G. Lenaers, S. Picaud, Cell Death Dis. 9 (2018) 287.
[5]X. Ouyang, J. Yang, Z. Hong, Y. Wu, Y. Xie, G. Wang, Biomed. Pharmacother. 130 (2020) 110577.
[6]W.T. Ham Jr, H.A. Mueller, D.H. Sliney, Nature 260 (1976) 153–155.
[7]D. van Norren, T.G.M.F. Gorgels, Photochem. Photobiol. 87 (2011) 747–753.
[8]I. Jaadane, P. Boulenguez, S. Chahory, S. Carré, M. Savoldelli, L. Jonet, F. Behar-Cohen, C. Martinsons, A. Torriglia, Free Radic. Biol. Med. 84 (2015) 373–384.
[9]Y.-M. Shang, G.-S. Wang, D. Sliney, C.-H. Yang, L.-L. Lee, Environ. Health Perspect. 122 (2014) 269–276.
[10]M. Schieber, N.S. Chandel, Curr. Biol. 24 (2014) R453–62.
[11]S. del Olmo-Aguado, A.G. Manso, N.N. Osborne, Photochem. Photobiol. 88 (2012) 1346–1355.
[12]J.-X. Tao, W.-C. Zhou, X.-G. Zhu, Oxid. Med. Cell. Longev. 2019 (2019) 6435364.
[13]J.R. Sparrow, J. Zhou, B. Cai, Invest. Ophthalmol. Vis. Sci. 44 (2003) 2245–2251.
[14]P. Chen, Z. Lai, Y. Wu, L. Xu, X. Cai, J. Qiu, P. Yang, M. Yang, P. Zhou, J. Zhuang, J. Ge, K. Yu, J. Zhuang, Cells 8 (2019).
[15]B.F. Godley, F.A. Shamsi, F.-Q. Liang, S.G. Jarrett, S. Davies, M. Boulton, J. Biol. Chem. 280 (2005) 21061–21066.
[16]N.N. Osborne, C. Núñez-Álvarez, S. Del Olmo-Aguado, Exp. Eye Res. 128 (2014) 8–14.
[17]T. Nakanishi-Ueda, H.J. Majima, K. Watanabe, T. Ueda, H.P. Indo, S. Suenaga, T. Hisamitsu, T. Ozawa, H. Yasuhara, R. Koide, Free Radic. Res. 47 (2013) 774–780.
[18]E.K. Ozkaya, G. Anderson, B. Dhillon, P.-O. Bagnaninchi, Exp. Eye Res. 189 (2019) 107817.
[19]N.D. Wangsa-Wirawan, R.A. Linsenmeier, Arch. Ophthalmol. 121 (2003) 547–557.
[20]M.T.T. Wong-Riley, Eye Brain 2 (2010) 99–116.
[21]H.S. Lee, L. Cui, Y. Li, J.S. Choi, J.-H. Choi, Z. Li, G.E. Kim, W. Choi, K.C. Yoon, PLoS One 11 (2016) e0161041.
[22]Y. Niwano, T. Kanno, A. Iwasawa, M. Ayaki, K. Tsubota, Br. J. Ophthalmol. 98 (2014) 990–992.
[23]V. Marek, S. Mélik-Parsadaniantz, T. Villette, F. Montoya, C. Baudouin, F. Brignole-Baudouin, A. Denoyer, Free Radic. Biol. Med. 126 (2018) 27–40.
[24]E.R. Gaillard, L. Zheng, J.C. Merriam, J. Dillon, Invest. Ophthalmol. Vis. Sci. 41 (2000) 1454–1459.
[25]A.J. Bron, G.F. Vrensen, J. Koretz, G. Maraini, J.J. Harding, Ophthalmologica 214 (2000) 86–104.
[26]L. Kessel, J.H. Lundeman, K. Herbst, T.V. Andersen, M. Larsen, J. Cataract Refract. Surg. 36 (2010) 308–312.
[27]M. Boulton, F. Docchio, P. Dayhaw-Barker, R. Ramponi, R. Cubeddu, Vision Res. 30 (1990) 1291–1303.
[28]R.H. Steinberg, Doc. Ophthalmol. 60 (1985) 327–346.
[29]M.M. LaVail, Invest. Ophthalmol. Vis. Sci. 19 (1980) 407–411.
[30]L. Feeney, Invest. Ophthalmol. Vis. Sci. 17 (1978) 583–600.
[31]C. Brandstetter, L.K.M. Mohr, E. Latz, F.G. Holz, T.U. Krohne, J. Mol. Med. 93 (2015) 905–916.
[32]J.R. Sparrow, K. Nakanishi, C.A. Parish, Invest. Ophthalmol. Vis. Sci. 41 (2000) 1981–1989.
[33]S.Y. Jeong, X. Gu, K.W. Jeong, Food Chem. Toxicol. 131 (2019) 110555.
[34]A. Alaimo, G.G. Liñares, J.M. Bujjamer, R.M. Gorojod, S.P. Alcon, J.H. Martínez, A. Baldessari, H.E. Grecco, M.L. Kotler, Arch. Toxicol. 93 (2019) 1401–1415.
[35]J. Loughman, P.A. Davison, J.M. Nolan, M.C. Akkali, S. Beatty, J. Optom. 3 (2010) 74–90.
[36]V.C. Lima, R.B. Rosen, M. Farah, Int J Retina Vitreous 2 (2016) 19.
[37]P.L. Pease, A.J. Adams, E. Nuccio, Vision Res. 27 (1987) 705–710.
[38]B.R. Hammond, L.M. Fletcher, F. Roos, J. Wittwer, W. Schalch, Invest. Ophthalmol. Vis. Sci. 55 (2014) 8583–8589.
[39]J.M. Stringham, P.V. Garcia, P.A. Smith, L.N. McLin, B.K. Foutch, Invest. Ophthalmol. Vis. Sci. 52 (2011) 7406–7415.
[40]J. Loughman, M.C. Akkali, S. Beatty, G. Scanlon, P.A. Davison, V. O’Dwyer, T. Cantwell, P. Major, J. Stack, J.M. Nolan, Vision Res. 50 (2010) 1249–1256.
[41]P. Bhosale, B. Serban, P.S. Bernstein, Arch. Biochem. Biophys. 483 (2009) 175–181.
[42]S.P. Sundelin, S.E. Nilsson, Free Radic. Biol. Med. 31 (2001) 217–225.
[43]P.S. Bernstein, B. Li, P.P. Vachali, A. Gorusupudi, R. Shyam, B.S. Henriksen, J.M. Nolan, Prog. Retin. Eye Res. 50 (2016) 34–66.
[44]A. Junghans, H. Sies, W. Stahl, Arch. Biochem. Biophys. 391 (2001) 160–164.
[45]A. Kijlstra, Y. Tian, E.R. Kelly, T.T.J.M. Berendschot, Prog. Retin. Eye Res. 31 (2012) 303–315.
[46]A.G. Murillo, S. Hu, M.L. Fernandez, Antioxidants (Basel) 8 (2019).
[47]E.J. Johnson, J.E. Maras, H.M. Rasmussen, K.L. Tucker, J. Am. Diet. Assoc. 110 (2010) 1357–1362.
[48]N.T. Stringham, J.M. Stringham, Perception 44 (2015) 1400–1415.
[49]B.R. Hammond Jr, E.J. Johnson, R.M. Russell, N.I. Krinsky, K.J. Yeum, R.B. Edwards, D.M. Snodderly, Invest. Ophthalmol. Vis. Sci. 38 (1997) 1795–1801.
[50]L. Ma, R. Liu, J.H. Du, T. Liu, S.S. Wu, X.H. Liu, Nutrients 8 (2016).
[51]A. Obana, Y. Gohto, R. Nakazawa, T. Moriyama, W. Gellermann, P.S. Bernstein, Sci. Rep. 10 (2020) 10262.
[52]N. Machida, M. Kosehira, N. Kitaichi, Nutrients 12 (2020).
[53]Y. Yao, Q.-H. Qiu, X.-W. Wu, Z.-Y. Cai, S. Xu, X.-Q. Liang, Nutrition 29 (2013) 958–964.
[54]J.M. Nolan, R. Power, J. Stringham, J. Dennison, J. Stack, D. Kelly, R. Moran, K.O. Akuffo, L. Corcoran, S. Beatty, Invest. Ophthalmol. Vis. Sci. 57 (2016) 3429–3439.
[55]J.M. Nolan, J. Loughman, M.C. Akkali, J. Stack, G. Scanlon, P. Davison, S. Beatty, Vision Res. 51 (2011) 459–469.
[56]L.M. Renzi-Hammond, E.R. Bovier, L.M. Fletcher, L.S. Miller, C.M. Mewborn, C.A. Lindbergh, J.H. Baxter, B.R. Hammond, Nutrients 9 (2017).
[57]E.R. Bovier, L.M. Renzi, B.R. Hammond, PLoS One 9 (2014) e108178.
[58]E.R. Bovier, B.R. Hammond, Arch. Biochem. Biophys. 572 (2015) 54–57.
[59]B.R. Hammond Jr, L.S. Miller, M.O. Bello, C.A. Lindbergh, C. Mewborn, L.M. Renzi-Hammond, Front. Aging Neurosci. 9 (2017) 254.
[60]J.M. Stringham, K.J. O’Brien, N.T. Stringham, Eye Vis (Lond) 3 (2016) 30.
[61]J.M. Stringham, N.T. Stringham, K.J. O’Brien, Foods 6 (2017).
[62]L. Ma, X.-M. Lin, Z.-Y. Zou, X.-R. Xu, Y. Li, R. Xu, Br. J. Nutr. 102 (2009) 186–190.
[63]J. Kvansakul, M. Rodriguez-Carmona, D.F. Edgar, F.M. Barker, W. Köpcke, W. Schalch, J.L. Barbur, Ophthalmic Physiol. Opt. 26 (2006) 362–371.
[64]R. Liu, T. Wang, B. Zhang, L. Qin, C. Wu, Q. Li, L. Ma, Invest. Ophthalmol. Vis. Sci. 56 (2014) 252–258.
[65]Age-Related Eye Disease Study 2 (AREDS2) Research Group, E.Y. Chew, T.E. Clemons, J.P. Sangiovanni, R.P. Danis, F.L. Ferris 3rd, M.J. Elman, A.N. Antoszyk, A.J. Ruby, D. Orth, S.B. Bressler, G.E. Fish, G.B. Hubbard, M.L. Klein, S.R. Chandra, B.A. Blodi, A. Domalpally, T. Friberg, W.T. Wong, P.J. Rosenfeld, E. Agrón, C.A. Toth, P.S. Bernstein, R.D. Sperduto, JAMA Ophthalmol. 132 (2014) 142–149.
[66]A. Yagi, K. Fujimoto, K. Michihiro, B. Goh, D. Tsi, H. Nagai, Appl. Ergon. 40 (2009) 1047–1054.
[67]T. Otsuka, M. Shimazawa, T. Nakanishi, Y. Ohno, Y. Inoue, K. Tsuruma, T. Ishibashi, H. Hara, J. Pharmacol. Sci. 123 (2013) 209–218.
[68]K. Ogawa, Y. Kuse, K. Tsuruma, S. Kobayashi, M. Shimazawa, H. Hara, BMC Complement. Altern. Med. 14 (2014) 120.
[69]E. Ooe, Y. Kuse, T. Yako, T. Sogon, S. Nakamura, H. Hara, M. Shimazawa, Mol. Vis. 24 (2018) 621–632.
[70]L. Tang, S. Bao, Y. Du, Z. Jiang, A.O. Wuliji, X. Ren, C. Zhang, H. Chu, L. Kong, H. Ma, Biomed. Pharmacother. 103 (2018) 829–837.
[71]J.-S. Chang, Y.-J. Lee, D.A. Wilkie, C.-T. Lin, J. Vet. Med. Sci. 80 (2018) 1108–1115.
[72]H. Pasantes-Morales, C. Cruz, Brain Res. 330 (1985) 154–157.
[73]W. Dayang, P. Dongbo, Cutan. Ocul. Toxicol. 37 (2018) 240–244.
[74]R. Maccarone, S. Di Marco, S. Bisti, Invest. Ophthalmol. Vis. Sci. 49 (2008) 1254–1261.
[75]F.D. Marco, S. Romeo, C. Nandasena, S. Purushothuman, C. Adams, S. Bisti, J. Stone, Am. J. Neurodegener. Dis. 2 (2013) 208–220.
[76]A. Laabich, G.P. Vissvesvaran, K.L. Lieu, K. Murata, T.E. McGinn, C.C. Manmoto, J.R. Sinclair, I. Karliga, D.W. Leung, A. Fawzi, R. Kubota, Invest. Ophthalmol. Vis. Sci. 47 (2006) 3156–3163.
[77]M. Yamauchi, K. Tsuruma, S. Imai, T. Nakanishi, N. Umigai, M. Shimazawa, H. Hara, Eur. J. Pharmacol. 650 (2011) 110–119.






No Comments Yet
Sign in or Register to Comment